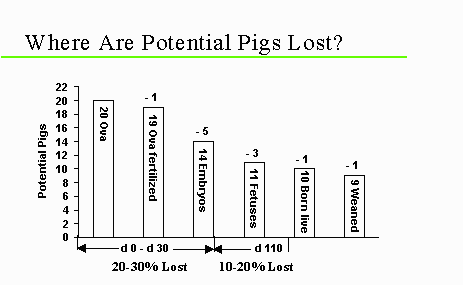
Purdue University 1998 Swine Day Report
B.A. Belstra, J.W. Frank, D.C. Kendall, B.T. Richert, M.A.
Diekman, and W.L. Singleton
Department of Animal Sciences
Approximately 30 to 50% of the ova ovulated do not result in a live piglet due to embryonic/fetal mortality during gestation in sows. Approximately 20 to 30% are lost during the first 30 days of gestation (early embryonic loss), with an additional 10 to 20% lost during the last 85 days (fetal loss) (Pope, 1994); see the figure below. The majority of early embryonic loss occurs during the period of blastocyst implantation (days 13 to 18), and this period is consequently of particular interest to researchers. Early embryonic loss is poorly understood, but a number of causes have been hypothesized. A portion of fetal loss may be due to the limited space for embryos in the uterus ("crowding"), and due to the limited capacity of the uterus to supply blood for nutrient exchange.
Early weaned sows often have a significantly increased level of early embryonic loss (Marstellar et al., 1996). Levels of progesterone (P4 ) and estrogens (particularly estradiol, E2 ) at specific periods during early gestation are critical for the development of the proper uterine environment for blastocyst implantation and for maintenance of pregnancy. Some evidence suggests early-weaned sows may have altered levels of P4 and E2 during early gestation (Varley et al., 1981; Varley et al., 1984). Other studies have found no effect of lactation length on steroid hormone profiles (Kirkwood et al., 1984). Treatment of sows with exogenous steroids during implantation has significantly increased the number of live pigs at farrowing (de Sa et al., 1981; Wildt et al., 1976).
The objectives of this study were to assess the impact of 1) lactation length and 2) exogenous steroid treatment on embryonic survival in the sow. We also wanted to 3) determine if the increased levels of embryonic mortality observed by other researchers in early-weaned sows were related to an altered steroid profile during early gestation.
Thirty-six second parity, high lean, European sows were either early weaned (EW; 10 to 15 day lactation; n=26) or conventionally weaned (CW; 26 to 37 day lactation; n=10). They were housed in an open front building with 12' x 32' pens in groups of 8 to 10. Sows were fed 5 lb/day of a 12.6% crude protein, 0.55% lysine gestation diet. Sows were checked for signs of estrus daily with a mature boar and were mated via AI to one of two boars at the first postweaning estrus. Inseminated sows were then housed individually in 8' x 10' pens and fed 5 lb of the same gestation diet daily from day 12 postmating until slaughter. Half of the sows from each weaning group were treated daily with 2 mL IM injections of a 25 mg progesterone and 1.25 ug estradiol-17b solution (HT; n=17) in the neck at 12:00 noon on days 14 to 20 postmating. The other half of each weaning group received a daily 2 mL IM injection of the carrier solution without the steroids as a control (C; n=17). Blood samples were collected via jugular venipuncture every other day beginning the day of weaning until days 25 to 26 of gestation, between 5:00 and 6:00 p.m. Serum concentrations of progesterone were determined by radioimmunoassay (estradiol-17b and estrone sulphate are currently being measured).
Sows were slaughtered at the Purdue abattoir in groups of 3 to 6 on days 28 to 30 of gestation. Reproductive tracts were quickly removed at slaughter and transported on ice to the laboratory. Uterine horns were separated at the bifurcation and the mesometrium was trimmed away from each horn. Uterine horns were opened by an incision along the length of each horn on the mesometrial side. Length of the uterine horn and spacing of the embryos within each horn was measured. Embryos were determined viable, moribund or dead by visual examination after removal from the chorioallantoic membranes. Crown-rump length (CRL) of each embryo was measured and embryos were weighed. Individual copora lutea (CL) were dissected from the ovaries to determine ovulation rate. The percentage of embryos present (% E) was calculated as [(No. of viable, moribund and dead embryos/No. of CL) x 100]. Similarly, the percent embryo survival (% ES) was calculated as [(No. of viable embryos/No. of CL) x 100].
Because there was no interaction between the factor levels in this 2 x 2 factorial design, only main effects will be presented (Table 1). Early-weaned sows tended to have longer wean to estrus intervals (WEI) than CW sows (5.5 vs. 4.6 days, P<.10). However, 5.5 days is still a relatively short WEI. The surprisingly short WEI interval of EW sows in this study may be the result of good lactation and postweaning sow management or may possibly be a characteristic of this particular genotype. There was a numerical trend for EW sows to have a decreased conception rate (70.8 vs. 90.0%) and an increased percentage of anestrus sows (7.7 vs. 0.0%) compared to CW sows. There were also more non-pregnant EW sows on day 30 due to cystic ovaries than in the CW treatment (16.6 vs. 0.0%), but this difference was not significant.
There appeared to be more non-pregnant HT sows (35.3 vs. 11.8%) and significantly more HT sows open due to cystic ovaries (23.5 vs. 0.0%, P<. 04) than C sows. However, it is unlikely that administration of the injection or the contents of the injection itself would cause termination of pregnancy or development of cystic ovaries. The steroids used should not have interfered with maternal recognition of pregnancy or implantation. Cystic ovaries usually develop postweaning when the estradiol produced by the developing follicles fails to induce an ovulatory LH surge from the pituitary. This is most common in sows from short lactations, because the hypothalamus is in a refractory feedback state and will not respond to elevated estradiol from the developing follicles. Cystic ovaries would probably have developed before the hormone treatment was initiated, around the time of estrus in other sows (3 to 8 days postweaning). There were no anestrus sows in the C or HT because only sows exhibiting estrus were inseminated and assigned to receive injections.
Ovulation rate was used as a covariate for the total number of embryos, % E and % ES. Even though there was a large range in ovulation rate among sows, mean ovulation rate was similar across treatments. The total number of embryos (viable + moribund + dead) tended to be greater in CW sows as compared to EW sows (16.9 vs. 13.5, P<.07), and the number of live embryos was significantly greater in CW sows than in EW sows (16.1 vs. 11.6, P<.02). Thus, the % E and % ES were significantly greater for CW sows as compared to EW sows (82.4 vs. 64.7%, P=.05, and 78.0 vs. 57.3%, P<.04, respectively). The HT tended to increase the total number of embryos present (16.7 vs. 13.6, P<.09) and the number of live embryos (15.3 vs. 12.3, P<.08) as compared to the C treatment. The % E and % ES for HT sows was numerically greater than for C sows (80.6 vs. 66.6%, P>.10, and 72.6 vs. 62.6%, P>.20, respectively). There was no interaction between lactation length and hormone treatment. Thus, HT essentially increased the numbers of viable embryos and % ES equally for both EW and CW sows (2.3 vs. 2.6 and 11.9 vs. 4.7%, respectively).
There were some differences in measurements of embryo growth and embryo spacing between treatments. However, these differences were due to the difference in the number of embryos present between treatments. When embryo growth parameters and embryo spacing were adjusted with a covariate, such as the number of viable embryos, treatment differences were lost. We chose to present the data unadjusted for numbers of embryos because it reflects the impact of the imposed treatments. Total weight of viable embryos was significantly greater for CW sows as compared to EW sows (28.0 vs. 18.8 g, P=.02), but mean viable embryo weight was not (1.77 vs. 1.61 g, P>.30). This difference in total embryo weight was simply due to the greater number of viable embryos present in CW sows. Total weight of viable embryos for HT sows was not greater than for C sows (25.3 vs. 21.8 g, P>.36), due to the smaller increase in the number of viable embryos for the HT sows as compared to the CW versus the EW sows. Crown-rump length, another measure of embryo growth, was not different between treatments. The amount of uterine space was significantly greater in C sows than HT sows (11.2 vs. 7.8 in, P=.05) and tended to be greater in EW sows than CW sows (10.9 vs. 8.1 in, P<.12). Again, this is a reflection of the lower number of viable embryos in EW and C sows. Variation in spacing of embryos was not different between treatments (P>.25).
Sows with short WEI of 3 to 5 days had significantly more viable embryos (14.6 vs. 10.8, P=.05) and tended to have a higher % ES (71.3 vs. 54.1%, P<.10) than sows with a longer WEI of 6 to 10 days (Table 2). Sows with short WEIs of 4 to 5 days have been shown to have a higher farrowing rate and larger litter size than sows with longer WEIs of 6 to 12 days by other researchers (Deckert et al., 1997; Wilson and Dewey, 1993). However, in this study, the difference in % ES between sows with short versus longer WEIs was not due to a difference in ovulation rate (20.3 vs. 19.7 ova, P>.70). The cause of this difference in % ES between sows with short and long WEIs has not been determined. However, it has become obvious that attention to management of sows during gestation and lactation can reduce the WEI and thus improve subsequent litter size.
These data highlight the impact of short lactations on sow rebreeding performance and embryonic survival. Decreasing lactation length from a relatively long lactation length often results in a greater number of pigs/sow/year. However, when a short lactation length is chosen (≤ 17 days), the incidence of sows exhibiting erratic estrus, prolonged estrus (cystic ovaries) or anestrus behavior increases and conception rate may be decreased. Different sow genotypes and production systems may be less prone to these reproductive problems, but very few can completely avoid them when short lactation lengths are implemented. Individual operations will need to determine their own ideal lactation length, because different nutrition and management programs have different abilities to balance the pig growth and pig output advantages of early weaning against the sow reproductive disadvantages.
Treatment with exogenous steroids during implantation may offer a moderate increase in number of embryos surviving to day 30 of gestation. However, our intent was not to develop a commercially feasible treatment, but to use exogenous steroid treatment as a model to study embryonic mortality in the sow. The effect of WEI on the number of viable embryos and % ES at day 30 suggests that management of sows before and during lactation to reduce the WEI would increase the number of pigs born alive at term. There are a number of management practices that can be implemented to improve embryo survival and litter size at term. However, it may take the utilization of several of them to minimize the loss in reproductive performance in the early-weaned sow.
Deckert, A.E., C.E. Dewey, J.J. Ford, and B.E. Straw. 1997. The influence of the weaning-to-breeding interval on ovulation rate in parity-two sows. Swine Health and Production 5:89-93.
de Sa, W.F., P. Pleumsamran, C.B. Morcom, and W.R. Dukelow. 1981. Exogenous steroid effects on litter size and early embryonic survival in swine. Theriogenology 15:245-255.
Kirkwood, R.N., K.R. Lapwood, W.C. Smith, and I.L. Anderson. 1984. Plasma concentrations of LH, prolactin oestradiol-17b and progesterone in sows weaned after lactation for 10 or 35 days. J. Reprod. Fert. 70:95-102.
Marsteller, T.A., G.A. Armbruster, D.B. Anderson, A.J. Wuethrich, J.L. Taylor, and J.T. Symanowski. 1996. Effect of lactation length on ovulation rate and embryo survival in swine. Swine Health and Production 5:49-56.
Pope, W.F. 1994. Embryonic mortality in swine. Embryonic Mortality in Domestic Animals. Edited by M.T. Zavy and R. D. Geisert. CRC Press, Boca Raton, FL pp. 53-77.
Varley, M.A., R.E. Peaker, and T. Atkinson. 1984. Effect of lactation length of the sow on plasma progesterone, oestradiol-17b and embryonic survival. Anim. Prod. 38:113-119.
Varley, M.A., T. Atkinson, and L.N. Ross. 1981. The effect of lactation length on the circulating concentrations of progesterone and oestradiol in the early weaned sow. Theriogenology 6:179-184.
Wildt, D.E., A.A. Culver, C.B. Morcom, and W.R. Dukelow. 1976. Effect of administration of progesterone and oestrogen on litter size in pigs. J. Reprod. Fert. 48:209-211.
Wilson, M.R., and C.E. Dewey. 1993. The associations between weaning-to-estrus interval and sow efficiency. Swine Health and Production 1:10-15.
Table 1. Effect of lactation length and exogenous steroid hormone treatment on sow rebreeding performance, embryo survival and embryo growth parameters.
Item |
EW |
CW |
Control |
Hormone |
ANOVAd |
|---|---|---|---|---|---|
No. of Sows |
26 |
10 |
17 |
17 |
|
Lactation Length, d |
13.0 ± 0.5 |
31.0 ± 0.8 |
22.2 ± 0.6 |
21.8 ± 0.6 |
LLa |
Sow Weaning Weight, lb |
472.5 ± 9.1 |
470.0 ± 14.1 |
470.2 ± 11.4 |
472.3 ± 11.4 |
ns |
Wean to Estrus Interval, d |
5.5 ± 0.2 |
4.6 ± 0.4 |
5.1 ± 0.3 |
4.8 ± 0.3 |
LLc |
% Pregnant Sows |
70.8% |
90.0% |
88.2% |
64.7% |
ns |
% Open Sows |
29.2% |
10.0% |
11.8% |
35.3% |
ns |
% Open, Cystic Ovaries |
16.6% |
0.0% |
0.0% |
23.5% |
HTb |
% Anestrus Sows |
7.7% |
0.0% |
|
|
ns |
Slaughter Live Weight, lb |
447.3 ± 8.2 |
462.4 ± 12.8 |
453.7 ± 10.3 |
455.9 ± 10.3 |
ns |
Gest. Day Slaughtered, d |
29.4 ± 0.2 |
29.3 ± 0.3 |
29.3 ± 0.3 |
29.4 ± 0.3 |
ns |
Ovulation Rate |
20.3 ± 0.9 |
20.8 ± 1.3 |
19.3 ± 1.0 |
21.8 ± 1.2 |
ns |
No. Total Embryos |
13.5 ± 1.0 |
16.9 ± 1.4 |
13.6 ± 1.1 |
16.7 ± 1.3 |
LLc,HTc |
No. Viable Embryos |
11.6 ± 1.0 |
16.1 ± 1.4 |
12.3 ± 1.1 |
15.3 ± 1.3 |
LLb,HTc |
% (Total Embryos/CL) |
64.7 ± 4.9 |
82.5 ± 6.9 |
66.6 ± 5.4 |
80.6 ± 6.5 |
LLb |
% (Viable Embryos/CL) |
57.3 ± 5.3 |
78.0 ± 7.5 |
62.6 ± 5.9 |
72.6 ± 7.1 |
LLb |
Total Viable Embryo Wt, g |
18.8 ± 2.2 |
28.0 ± 3.0 |
21.8 ± 2.4 |
25.0 ± 2.7 |
LLb |
Mean Viable Embryo Wt, g |
1.61 ± 0.09 |
1.77 ± 0.12 |
1.70 ± 0.10 |
1.68 ± 0.11 |
ns |
Embryo CRL, in |
0.94 ± 0.03 |
1.00 ± 0.04 |
0.97 ± 0.04 |
0.96 ± 0.04 |
ns |
Uterine Space/Embryo, in |
10.9 ± 1.0 |
8.1 ± 1.4 |
11.2 ± 1.1 |
7.8 ± 1.3 |
LLc,HTb |
a These effects were significant at P<.01.
b These effects were significant at P<.05.
c These effects were trends at P<.10.
d LL = effect of lactation length: early weaned (EW) vs.
conventionally weaned (CW).
HT = effect of hormone treatment: control vs. hormone.
Table 2. Effect of wean to estrus interval on the ovulation rate, number of viable embryos and the % embryo survivala at day 30 of gestation in sows.
Wean to Estrus Interval |
No. Sows |
Ovulation Rate |
No. Viable Embryos |
% Embryo Survival |
|---|---|---|---|---|
3-5 days |
20 |
20.3 ± 0.9 |
14.6 ± 0.9 |
71.3 ± 4.8 |
6-10 days |
7 |
19.7 ± 1.5 |
10.8 ± 1.7 |
54.1 ± 8.9 |
ANOVA |
|
ns |
P=.05 |
P<.10 |
a % embryo survival = (No. viable embryos/No. of CL) x 100.
Index of 1998 Purdue Swine Day Articles
If you have trouble accessing this page because of a disability, please email anscweb@purdue.edu.