Purdue University 1996 Swine Day Report
A.L. Bollinger, M.E. Wilson, A.P. Pusateri, and M.A. Diekman
Department of Animal Sciences
Photoperiodism has been studied extensively in many mammals. It has been linked to reproduction in seasonal breeders such as sheep, horses, hamsters, ferrets, deer, and mink. The cue that seasonally breeding animals must receive to initiate or continue reproduction activity is received by the eye. This message is transferred from the eye to the pineal gland, the internal clock that regulates the synthesis and release of the hormone melatonin. The relationship between melatonin secretion and seasonality has been studied to some extent in pigs but much of the evidence is conflicting. Researchers in Australia report that "in pigs, as in other species, the concentration of melatonin in plasma increases in the dark and the duration of the nocturnal increase depends on photoperiod." In 1993, our laboratory demonstrated that only 30% of 3, 4, 5, and 6 month old prepubertal (non-cycling) gilts showed a 2-fold increase in melatonin during darkness. From these data, Mike Green determined that "the small nocturnal elevation coupled with the highly variable response suggests the lack of a timekeeping role for melatonin in the domestic pig." The objective of the study reported here was to determine if a nocturnal rise in serum melatonin was necessary before a gilt can attain puberty.
Twenty crossbred gilts (Yorkshire x Hampshire x Duroc) were transported from a conventional, mechanically-ventillated, grow-finish swine unit at 5 months of age (approximately 216 ± 9.3 lb) to an environmentally-controlled building maintained at 18C. Gilts were penned in groups of 10 under an artificial light regimen of 12:12 light:dark and allowed to acclimate to the facility for 30 days. Lighting was supplied by 250W incandescent bulbs and controlled by timers which turned the lights on at 0700 h and off at 1900 h. Light intensity at eye level to the gilts measured approximately 700 lux. A 16% crude protein ration was available to the pigs ad libitum until they were transported to the experimental housing unit; a 14% crude protein ration was fed for the remainder of the experiment. Three mature ewes were transported at the same time to serve as controls for the environmental conditions. Food and water were available ad libitum for the ewes throughout the experiment.
At the end of the acclimation period, a blood sample was drawn from each gilt and assayed for progesterone (P4) to determine reproductive status. Gilts were determined to be pubertal if serum concentrations of P4 were greater than 500 pg/ml and were eliminated from the selection pool. Gilts were weighed and the two lightest and the two heaviest gilts were eliminated as well. From the remaining group, 10 gilts (309 ± 12.8 lb) were randomly chosen and individually penned for the remainder of the experiment.
Blood samples were drawn from surgically implanted jugular catheters according to the experimental protocol outlined in Table 1. The experiment was repeated in its entirety two additional times for a total of 30 gilts. Serum concentrations were quantified by radioimmunoassay.
Serum progesterone concentrations were measured for the gilts throughout the experiment in order to determine the time of ovulation and subsequently, the first day of the estrous cycle. Serum concentrations of progesterone for pubertal gilts in all three replications rose significantly after the onset of puberty while serum concentrations of progesterone for gilts that did not exhibit signs of estrus remained at baseline.
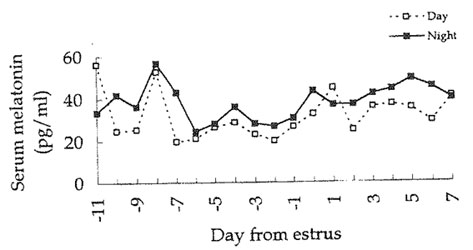
Serum concentrations of melatonin were similar (P>0.05) between pubertal and prepubertal gilts during light and dark periods in all three replications. Serum melatonin concentrations fluctuated considerably during the day and the night within each pig as well as within each replication. Data were standardized to day at estrus (day 0) for the pubertal gilts, and means were calculated according to the number of gilts on each day (day -11 through 7 from estrus). Resulting melatonin concentrations are shown in Figure 1. The number of nocturnal increases per number of bleeding periods for all gilts and ewes in all three replications is outlined in Table 2. Only 65 of 406 bleeding periods in the gilts (16%) exhibited a nocturnal rise in serum melatonin. The majority of bleeding periods in the ewes (85.1%) demonstrated nocturnal rises in melatonin throughout the experiment.
Data clearly indicate that the natural nocturnal increases in serum melatonin did not change or increase as gilts approached or attained puberty when housed under a 12L:12D lighting regimen. The lack of a nocturnal rise in serum melatonin in prepubertal gilts as they reach puberty is not surprising when compared to previous work done by our lab. As of yet, there has been no consistency in the reports on melatonin in pigs. The conclusions of Paterson and colleagues have been in direct contrast to those of this laboratory. They have boldly stated that the pig is a short day breeder and reproduction is regulated in the pig according to melatonin released from the pineal gland. There may be various reasons for the discrepancies. Though both laboratories are using pigs of common ancestry it is possible that the genetics of the Australian herds have changed enough to cause significant differences in their reactions to light and their release and synthesis of melatonin. Methods of production also may have had some effect as a majority of pigs in the US are raised in environmentally-controlled conditions, whereas Australian pigs are often raised outdoors with year-round exposure to external environmental conditions.
Melatonin secretory patterns for the sheep illustrate consistent nocturnal increases in relation to baseline daytime concentrations. Thus, the environmental conditions excluding photoperiod may be ruled out as an explanation for the lack of nocturnal rises in the gilts at, before or after puberty. The number of nocturnal rises were not changed in gilts that attained puberty and those that did not. The fact that some of the gilts did show nocturnal increases in serum melatonin that were not associated with estrus implies that a nocturnal increase in serum melatonin is not necessary for a gilt to attain puberty.
Table 1. Schedule for sampling, surgeries, and boar exposures.
Day -30
Gilts transported to environmentally controlled room and allowed to acclimate for 30 days.
Day 0
Progesterone screening and weighing of gilts.
Day 1
Surgical implantation of venal catheters on 5 of 10 gilts.
Day 2
Surgical implantation of jugular vein catheters on second 5 gilts.
Day 3
First sampling day for all 10 gilts.
| Sampling Schedules | |
Day |
1100 |
Day |
1130 |
Day |
1200 |
Night |
2300 |
Night |
2330 |
Night |
2400 |
Day 5
All gilts driven around in truck and exposure to the boar was initiated.
Day 6
Boar exposures every morning for the duration of the experiment for 20 minutes.
| Sampling Schedules | |
Day |
1100 |
Day |
1130 |
Day |
1200 |
Night |
2300 |
Night |
2330 |
Night |
2400 |
Day 12
Any gilts that had not showed estrous were removed from the experiment.
Gilts in estrus were bled until 7 days following the onset of estrus.
Table 2.Number and percentage of nocturnal rises per bleeding period for ewes and gilts.
Item |
Ewes |
Gilts |
|---|---|---|
Nocturnal Rises, N - Rep I |
26/39 |
14/131 |
Nocturnal Rises, N - Rep II |
30/32 |
23/152 |
Nocturnal Rises, N - Rep III |
30/30 |
28/123 |
Nocturnal Rises, N |
86/101 |
65/406 |
Nocturnal Rises, % |
85.1 |
16 |
Figure 1. Serum concentrations of melatonin in gilts before and after attainment of puberty during daytime and nighttime.
Index of 1996 Purdue Swine Day Articles
If you have trouble accessing this page because of a disability, please email anscweb@purdue.edu.